Peraturan sintésis koléstérol
Peraturan sintésis koléstérol - énzim konci (HMG-CoA reductase) dilakukeun ku sababaraha cara.
Fosforilasi / dephosphorylasi HMG réduktase. Kalayan paningkatan dina rasio insulin / glukagon, fosforus template énzim ieu pas kana kaayaan anu aktip. Tumindak insulin dilaksanakeun ngalangkungan 2 énzim.
HMG-CoA réduktase kinase fosfatase, anu kéngingkeun kinase janten kaayaan dephosforilasi anu henteu aktip:
Fosfasease HMG-CoA réduktase ku ngarobih kana kaayaan aktif dephosphorylated. Hasil tina réaksi ieu mangrupikeun formasi bentuk aktif dephosforus HMG-CoA réduktase.
Akibatna, dina nalika nyerep, koléstérér naék. Salila periode ieu, kasadiaan substrat awal pikeun sintésis koléstérol - acetyl - CoA nambahan (akibat tina tuangeun tuangeun tuangeun karbohidrat sareng lemak, saprak CoA acetyl kabentuk nalika ngarecah glukosa sareng asam lemak).
Dina kaayaan postabsorbent, glukagon liwat proteingenase A ngrangsang fosforilasi HMG - CoA - réduktase, balik janten kaayaan anu teu aktip. Padamelan ieu ditingkatkeun ku kanyataan yén dina waktos anu sami, glukagon merangsang fosforilasi sareng teu aktivasi fosfatase HMG-CoA réduktase, ku kituna ngajaga HMG-CoA réduktase dina kaayaan aktif fosforil. Hasilna, sintésis koléstérol dina periode postabsorption sareng salami puasa dicegah ku sintésis endogen. Lamun eusi koléstérol dina dahareun dibawa ka 2%, maka sintésis kolégérogén endogenous menurun. Tapi gencatan sintésis koléstérol lengkep henteu lumangsung.
Gelar ngahambat biosintésis kolésteris dina pangaruh koléstérol anu asalna tina dahareun bénten-béda ti jalma ka jalma. Ieu nunjukkeun personalitas tina prosés formasi koléstérol. Ku ngukur intensitas sintésis kolésterol, mungkin waé ngirangan konsentrasi dina getih.
Lamun kasaimbangan antara asupan koléstérol sareng dahareun sareng sintésisna dina awak dina hiji sisi sareng ékskrési asam bili na koléstéris lain rusak, konsentrasi koléstérol dina jaringan sareng parobihan getih. Akibat anu paling serius pakait sareng paningkatan konsentrasi koléstérol getih (hypercholesterolemia), sedengkeun kamungkinan ngembangkeun aterosklerosis sareng kolelithiasis naek.
Hiperkolesterolemia Familial (HCS) - formulir ieu langkung umum - sakitar 1 pasien per 200 urang. Cacat anu diwariskeun dina HCS nyaéta palanggaran nyerep LDL ku sél, sareng ku sabab panurunan dina katabolisme LDL. Salaku hasilna, konsentrasi LDL dina getih naék, ogé koléstérol saprak aya pisan di LDL. Kukituna, sareng HCS, déposisi koléstérol dina jaringan, hususna dina kulit (xanthomas), dina dinding arteri mangrupikeun ciri-ciri.
Inhibition sintésis HMG-CoA réduktase
Produk ahir jalur métabolik koléstérol. Éta ngurangan laju transkripsi gén HMG-CoA réduktase gén, sahingga ngahambat sintésisna nyalira. Ati aktip nyintésis asam bili tina koléstérol, sareng ku kituna asam bili ngahambat kagiatan gén Hédg-CoA. Kusabab HMG-CoA réduktase aya saatos sintésis sakitar 3, panyambutan sintésis énzim koléstérol ieu nyaéta pangaturan anu épéktip.
Teu mendakan naon anu dipilari? Anggo pamilarian:
Bursa éster kolesterol
Dana koléstérol ngandung éster koléstérol sareng éster kolesterol bébas, anu dipendakan na sél na lipoprotein getih.
Bagéan II. Métabolisme sareng énergi
Dina sél, éstérifikasi koléstérol lumangsung sareng aksi acyl-CoA-kolésterol-acyltransferase (AChAT):
Acyl-CoA + Kolesterol - * HS-KoA + Acylcholesterol
Dina sél manusa, linoleylcholesterol utamana kabentuk. Beda sareng koléstérol bébas, éster na mémbran sél dikandung dina jumlah sakedik sareng dipendakan biasana dina sitosol salaku bagian tina tetes lipid. Wangunan éster tiasa dianggap, dina hiji sisi, salaku mékanisme pikeun ngaleungitkeun koléstérol tina mémbran, sareng sanésna, salaku mékanisme pikeun neundeun koléstérol dina sél. Mobilisasi sumber lumangsung kalayan partisipasi énzim ésterase anu nghidrolkeun éster koléstérol:
Acylcholesterol + H 2O - * asam lemak + kolesterol
Sintésis jeung hidrolisis éster lumangsung dina seueur sél, tapi hususna aktip dina sél korteks adrenal: dina sél ieu dugi ka 80% sadaya koléstéris diwakilan ku éster, sedengkeun dina sél séjén biasana kurang tina 20%.
Wangunan éster dina liponroteins getih lumangsung sareng partisipasi lecithin-kolesterol acyltransferase (LHAT), anu ngatalisan transfer sésana acyl tina i-posisi lésitin ka koléstérol (Gbr. 10,31). LHAT dibentuk dina ati, disumputkeun kana getih sareng meungkeut lipoproteins. Tingkat éstérifikasi pikeun lipoprotein anu béda-béda béda sareng gumantung ayana apolipoprotein anu ngaktifkeun LHAT (utamina apo-AT, ogé CI) atanapi ngahambat (C-II) enzim ieu. LHAT paling aktif dina HDL, dimana apo-AT kedah olee 2/3 sadaya protéin. honors coli Masalah Panggedéna kabentuk éster asam oleat na linoléat. The lipoproteins séjén éster formasi lumangsung dina laju laun ti di HDL.
Gbr. 10.31. Wangunan éster koléstérol dina kaayaan LHAT
LHAT dilokalisasi dina lapisan permukaan HDL sareng nganggo koléstérol dina monolayer fosfatip janten substrat. Éster ésterolol dibentuk di dieu, kusabab lengkep hidrofobicity, kirang disimpen
Bab 10. Metabolisme sareng Fungsi Lipid
fosfolipid monolayer sareng kacemplung dina inti lipid lipoprotein. Dina waktu anu sarua, tempat pikeun koléstérol dibebaskeun dina monolayer fosfolipid, anu tiasa dieusi koléstérol tina mémbran sél atanapi tina lipoprotein sanés. Kituna, HDL katingalina mangrupikeun perangkap koléstérol salaku hasil tina LHAT.
Sintésis Asam bili
Dina ati, bagian koléstérol dirobah jadi asam bili. Asam Gallik tiasa dianggap salaku turunan asam cholanic (Gbr. 10,32).
Asam Cholanic sapertos kieu teu kabentuk dina awak. Dina hépatosit, koléstérol langsung ngahasilkeun asam chenodeoxycholic and cholic - asam bili primér (Gbr. 10.33, tempo ogé Gbr. 10,12).
Biosintésis kolesterol
Biosintésis koléstérol lumangsung dina reticulum endoplasmic. Sumber sadaya atom karbon dina molekul nyaéta acetyl-SCoA, anu asalna ti mitochondria salaku bagian citrate, sakumaha sintésis asam lemak. Biosintésis koléron meakeun 18 molekul ATP sareng 13 molekul NADPH.
Wangunan koléstérol lumangsung dina leuwih ti 30 réaksi, anu tiasa dikelompokkeun dina sababaraha hambalan.
1. Sintésis asam mevalonik
Dua réaksi sintésis munggaran saluyu sareng réaksi ketogenesis, tapi saatos sintésis 3-hydroxy-3-methylglutaryl-ScoA, énzim ngalebetkeun hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA réduktase), ngabentuk asam mevalonik.
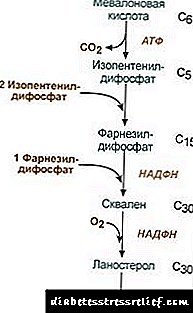 |
Skéma réaksi sintésis kolesterol2. Sintésis isopentenyl diphosphate. Dina tahap ieu, tilu résidu fosfat meungkeut asam mevalonik, teras éta déskriptil sareng dehidrogenasi. 3. Saatos ngahijikeun tilu molekul isopentenyl diphosphate, farnesyl diphosphate disintésis. 4. Sintésis squalene lumangsung nalika dua résidu farnesyl diphosphate kaiket. 5. Saatos réaksi anu rumit, liniér squalene ngéngingkeun ka lanosterol. 6. Ngaleungitkeun kalolobaan kelompok metil, pamulihan sareng isomerisasi molekul ngabalukarkeun penampilan koléstérol. Peraturan kagiatan hydroxymethylglutaryl-S-CoA réduktase3. Tingkat biosintésis koléstérol ogé gumantung kana konsentrasi protein pamawa khususnyayogikeun pikeun ngariung sareng angkutan metabolit sintésis panengah hidrofobik. Anjeun tiasa naros atanapi ngantepkeun pendapat anjeun. Titik aturan utama nyaéta réaksi pembentukan asam mevalonik. 1. Peraturan Allosteric. Kolesterol, sareng dina ati - sareng asam bili ngahambat réduktase HMG-CoA. 2. Représis sintésis HMG-CoA kolésterérasi réduktase. 3. Peraturan ku fosforilasi-dephosforilasi HMG-CoA réduktase, bentuk non-fosforilasi aktif. Glucagon nyababkeun deaktivasi, sareng insulin nyababkeun aktivasina ngaliwatan réaksi réaksi anu kompleks. Maka, laju sintésis kolésterol parobihan ku parobahan dina nyerep sareng postabsorption. 4. Laju sintésis dina réduksi HMG-CoA dina ati tunduk kana turunna diurnal anu jelas: maksimal tengah wengi, sareng minimum dina subuh. Bursa éster kolesterol Dina sél Éstérifikasi koléstérol lumangsung nalika kakeunaan acyl-CoA-koléstér-acyltransferase (AHAT): Acyl-CoA + Cholesterol ® NS-CoA + Acylcholesterol Dina sél, utamina linoleylcholesterol dibentuk. Éster kapanggih utamana dina sitosol salaku sabagian titisan lipid. Wangunan éster tiasa dianggap, dina hiji sisi, salaku mékanisme pikeun ngaleungitkeun koléstérol tina mémbran, sareng sanésna, salaku mékanisme pikeun nyimpen koléstérol dina sél. Mobilisasi cadangan lumangsung sareng partisipasi énzim ésterésterololisis éster kolesterol: Acylcholesterol + N2O® Asam Lemak + Koléstérol Sintésis jeung hidrolisis éster utamina aktip dina sél korteks adrénal. Dina lipoproteins getih formasi ester lumangsung sareng partisipasi na lecithin-kolesterol-acyltransferase (LHAT), ngatalisan transisi sésa acyl tina lésitin ka koléstérol. LHAT kabentuk dina haté, disékrésikeun kana aliran getih sareng napel dina lipoprotein. LHAT paling aktif dina HDL, dimana dilokalisasi dina lapisan permukaan. Éster koléster anu dibentuk di dieu mangrupikeun hidrofobik sareng nyumput dina inti lipid. Dina monolayer fosfolipid, aya rohangan bébas koléstérol, anu tiasa dieusi koléstérol tina mémbran sél atanapi tina lipoprotein sanés. Kituna, HDL katingalina mangrupikeun perangkap koléstérol salaku hasil tina LHAT. Sintésis Asam bili Dina ati, bagian koléstérol dirobah jadi asam bili. Asam bili tiasa dianggap salaku turunan asam cholanic. Asam Cholanic sapertos kieu teu kabentuk dina awak. Dina hépatosit tina koléstérol, asam bili primér kabentuk - chenodeoxycholic jeung kritis. Saatos disékrésikeun bili kana peujit dina aksi énzim tina flora peujit ti aranjeunna asam empedu sekundér kabentuk - litiokolik jeung deoxycholic. Éta diserep tina peujit, kalayan getih urat portal asupkeun ati, teras kana bili. Bile ngandung asam asam tumpukan utamina, s.a., sanyawana nganggo glikogén atanapi taurine. Konsentrasi asam bili dina bili kira-kira 1%. Bagian utama asam biliin aub sirkulasi hépatoenterik.Bagéan leutik asam bili - sakitar 0,5 g per dinten - dikaluarkeun dina na. Ieu diimbangan ku sintésis asam bili anyar dina ati, dana asam bili diropéa sakitar 10 dinten. Kolesterol ogé dikaluarkeun utamana tina peujit. Éta asup kana peujit dina dahareun sareng tina ati janten bagian tina bili. Kolesterol diserep kana getih ngandung fraksi anu asalna tina bili (kolesterol endogendisintésis dina ati), sareng fraksi na diturunkeun tina dahareun (koléstérol exogenous) Ngaleungitkeun koléstérol tina jaringan lumangsung ku oksidasi na asam bili dina ati, dituturkeun ku ékskrésina ku na (kira-kira 0,5 g per dinten) sareng ékskrési kolesterol anu teu dirobih (ogé sareng tai). Dina kaayaan stasion: (Kolesteroltungtungna + Koléstérolmantan) - (Koléstéroljalanna + Asam bilijalanna) = 0 Upami kasaimbangan ieu kaganggu, konsentrasi koléstérol dina jaringan sareng dina getih robah. Ngaronjat koléter getih - hypercholesterolemia. Ieu ningkatkeun probabilitas atherosclerosis sareng panyakit batu. PERATURAN LIPID Métabolisme lipid diatur ku sistem saraf pusat. Lila pisan setrés émosional négatip, paningkatan dina sékrési catecholamines kana aliran getih tiasa nyababkeun kaleungitan beurat. Laku lampah glukagon dina sistem lipolitik sami sareng tindakan katekolamin. Adrenalin jeung norepinephrine ningkatkeun kagiatan jaringan lipase sareng laju lipolisis dina jaringan adipose, salaku hasilna, eusi asam lemak dina plasma getih naék. Insulin boga pangaruh sabalikna ti adrenalin sareng glukagon dina lipolysis sareng mobilisasi asam lemak. Tumuwuh hormon ngarangsang lipolisis, nyorong sintésis acenylate siklik. Hipofunction hipofisis ngabalukarkeun déposisi lemak dina awak (obesitas hipofisis). Thyroxine, hormon seksogé mangaruhan métabolisme lipid. Ngaleungitkeun kelenjar kelamin dina sato nyababkeun déposisi lemak kaleuwihan. PITIK METABOLIK LIPID Kolesterol mangrupakeun stéroid khusus pikeun organisme sato. Tempat utama kabentukna dina awak manusa nyaéta ati, dimana 50% koléstéris disintésis, 15-20% kabentuk dina usus leutik, sesa disintésis dina kulit, korteks adrenal, sareng gonads. Sumber sumber pambentukan dana koléstérol sareng cara pengeluaran na ditepikeun dina Gbr. 22.1. Gbr. 22.1. Wangunan jeung distribusi koléstérol dina awak. Koléstérasi awak manusa (total kira-kira 140 g) sacara kondisyon tiasa kabagi kana tilu kolam: 30 g), gancang séukeur, diwangun ku koléstérol peujit, plasma getih, ati sareng organ parenchymal sanés, pembaruan kajantenan dina 30 dinten (1 g / dinten). 50 g), lalaunan pertukaran koléstérol organ sareng jaringan sanés, 60 g), koléstérol anu saleresna disilihtukeurkeun tina tulang tonggong sareng uteuk, jaringan konéktip, laju update diitung sababaraha taun. Sintésis koléstérol lumangsung dina sitosol sél. Ieu mangrupikeun salah sahiji jalur métabolik anu paling panjang dina awak manusa. Éta ngaluarkeun 3 hambalan: mimiti ditungtungan ku pembentukan asam mevalonik, anu kadua sareng formasi squalene (struktur hidrokarbon linier anu diwangun ku 30 atom karbon). Dina tahap katilu, squalene dirobih janten molekul lanosterol, teras aya 20 réaksi berturut-turut anu ngarobah lanosterol janten koléstérol. Dina sababaraha jaringan, gugus koléstérol hidroksil ngahasilkeun éster. Réaksi dikandung ku énzim intracellular AHAT (acylCoA: koléstil acyltransferase). Réaksi éstérifikasi ogé lumangsung dina getih dina HDL, dimana énzim LHAT perenahna (lecithin: koléstiltransferase). Éster koléstérol nyaéta bentuk dimana diangkut ku getih atanapi disimpen dina sél. Dina getih, kira-kira 75% koléstérol aya dina bentuk éster. Sintésis kolesterol diatur ku mangaruhan kagiatan sareng jumlah énzim penting tina prosés - 3-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA reductase). Ieu kahontal ku dua cara: 1. Fosforilasi / dephosphorylasi HMG-CoA réduktase. Insulin ngrangsang dephosforilasi tina HMG-CoA réduktase, teras narjamahkeun kana kaayaan anu aktip. Ku sabab kitu, dina periode nyerep, sintésis koléstéris naek. Salila periode ieu, kasadiaan substrat mimiti sintésis, acetyl-CoA, ogé nambahan. Glucagon gaduh pangaruh tibalik: ngalangkungan protéin kinase A, ngarangsang fosforilasi réduktase HMG-CoA, ngarobah kana kaayaan anu teu aktip. Hasilna, sintésis koléstérol dina periode postabsorption sareng salami puasa dipeungpeuk. 2. Tirasi sintésis HMG-CoA réduktase.Kolesterol (produk akhir jalur métabolik) ngirangan laju transkripsi gén HMG-CoA réduktase gén, sahingga ngahambat sintésisna nyalira, sareng asam bili ogé ngabalukarkeun pangaruh anu sami. Pengangkut kolesterol getih dilaksanakeun minangka bagian tina ubar. LPs nyayogikeun koléstérén exogenous dina jaringan, nangtoskeun aliran antara organ sareng ékskrési tina awak. Koléstérol éksina dikirimkeun ka ati janten bagian tina ChM sésa. Aya, bareng jeung koléstérol endogen anu disintésis, éta ngabentuk dana umum. Dina hépatosit, TAG sareng koléstérér dibungkus dina VLDL, sareng dina bentuk ieu disékrésikeun kana getih. Dina pangaruh LP-lipase, nghidrolisis TAG ka gliserol sareng asam lemak dina getih, VLDLPs mimiti dirobih kana STLP teras ka LDLPs anu ngandung dugi ka 55% koléstérol sareng éster na. LDL mangrupikeun bentuk transportasi koléstérol utama dimana dikirimkeun ka jaringan (70% koléstérol sareng éster na getih nyaéta bagian tina LDL). LDL tina getih asup kana ati (nepi ka 75%) sareng jaringan sanésna anu ngagaduhan reséptor LDL dina permukaanna. Lamun jumlah koléstérol nu ngalebetkeun sél ngaleuwihan kabutuhanna, maka sintésis reséptor LDL diteken, anu ngirangan aliran koléstérol tina getih. Kalayan panurunan dina konsentrasi koléstérol bébas dina sél, sabalikna, sintésis reséptor diaktipkeun. Hormone aub dina pangaturan sintésis reséptor LDL: insulin, triiodothyronine sareng hormon seks ningkatkeun formasi reséptor, sareng turunna glukokortikoid. Dina anu disebut "angkutan balik koléstérol", i. jalur anu ngajamin kasumpingan koléstérol kana ati, HDL maénkeun peran utama. Éta disintésis dina ati dina bentuk prékursor immature anu praktis henteu ngandung koléstérol sareng TAG. HDL prékursor dina getih jenuh koléstérol, nampi tina pirs jeung mémbran sél anu sanés. Mindahkeun koléstérol ka HDL ngalibatkeun énzim LHAT anu aya dina tempatna. Énzim ieu nampeu résidu asam lemak tina fosfatidilch (lecithin) ka koléstérol. Hasilna, molekul hidrofobik éster koléstér kabentuk, anu gerak dina jero HDL. Ku kituna, henteu diinum HDL, diperkaya ku koléstérol, janten HDL 3 - partikel dewasa sareng ageung. HDL 3 éster kolesterol bursa pikeun TAG anu dikandung dina VLDL sareng STD sareng partisipasi protéin khusus anu mindahkeun éster koléter antara lipoproteins. Dina hal ieu, HDL 3 balikkeun kana HDL2, anu ukuranna ningkat kusabab akumulasi TAG. VLDL sareng STDL dina pangaruh LP-lipase dirobih kana LDL, anu biasana ngahasilkeun koléstérol na ati. Bagéan leutik koléstérol dikirim ka ati HDL2 sareng HDL. Sintésis asam bili. Dina ati, 500-700 mg asam bilium unggal dinten disintésis tina koléstérol. Pembentukanna kalebet réaksi pangenalan gugus hidroksil kalayan partisipasi hidroksilase sareng réaksi oksidasi parsial rantay sisi koléstér (Gbr. 22.2): Gbr. 22.2. Skéma pembentukan asam bili. Réaksi sintésis munggaran - formasi 7-a-hydroxycholesterol - nyaéta pangaturan. Aktivitas énzim anu ngatalisan réaksi ieu dipeungpeuk ku produk ahir tina jalur, asam bili. Mekanisme pangaturan anu séjén nyaéta fosforilasi / dephosforilasi énzim (bentuk fosforilasi 7-a-hydroxylase aktip). Perda ogé tiasa dilaksanakeun ku cara ngarobah jumlah énzim: koléstérol nyababkeun transkripsi gén 7-a-hydroxylase, sareng asam tambang. Hormon tiroid nyurung sintésis 7-a-hydroxylase, sareng panindangan estrogen. Pangaruh sapertos éstrogén dina sintésis asam bili na ngajelaskeun naha panyakit batu permata lumangsung dina awéwé 3-4 kali langkung sering dibanding lalaki. Asam kolik jeung chenodeoxycholic dibentuk tina koléstérum disebut "asam bili primér". Sebagéan ageung asem ieu ngalaman konjugasi - ditambah molekul glikin atanapi taurine ka gugus karboksil asam bili. Konjugasi dimimitian ku formasi bentuk asam bili - turunan CoA, teras taurine atanapi glikol dipasang, sareng salaku hasilna 4 varian konjugér dibentuk: taurocholic and taurohenodeoxycholic, glycocholic and glycohenodeoxycholic acid. Émulsifiers sacara signifikan tibatan asam bili asli. Konflik sareng glikol dibentuk 3 kali langkung seueur ti taurine, sabab jumlah taurine dina awak dibatesan. Dina peujit, sajeroning sajeroning sajeroning asam urat primér dina kaayaan énzim baktéri dirobih jadi asam bili sekundér. Asam Deoksiokik, dibentuk tina kritik, sareng litiokolik, dibentuk tina deoxycholic, henteu larut sareng langkung kaserep dina peujit. Sakitar 95% asam urat anu asupkeun peujit kana ati ngalangkungan urat portal, teras disékrésikeun deui kana bili na dianggo deui dina émosiifikasi lemak. Ieu jalur asam bili disebut sirkulasi enterohepatic. Kalayan najis, asam bili sekundér biasana dipiceun. Panyakit gallstone (cholelithiasis) mangrupikeun prosés patologis dimana batu ngabentuk dina hampru, dasarna nyaéta koléstérol. Pelepasan koléstérol kana bili na kedah dibarengan ku sékrési asam bili na fosforipid anu ngajaga molekul koléstérol hidrofobik dina kaayaan micellar. Alesan anu ngarah kana parobahan dina rél asam bili na koléstérol dina bili: katuangan beunghar koléstérol, nutrisi kalori tinggi, stagnasi bili dina hampru, sirkulasi enterohepatic gangguan, sintésis gangguan tina asam bili, inféksi hampru. Di sabagéan ageung pasién anu ngandung kolelithiasis, sintésis koléstéris naék, sareng sintésis asam bili ti dinya ngalambatkeun, anu nyababkeun disproporasi dina jumlah koléstil sareng asam biliin dina kana tihang. Hasilna, koléstérér mimiti mendakan dina hampru, ngabentuk mendakan kentel anu laun bakal hese. Kadang-kadang ditingali sareng bilirubin, protéin sareng uyah kalsium. Batu ukur kalebet ukur koléstérol (batu koléstérol) atanapi campuran koléstérol, bilirubin, protéin sareng kalsium. Batu kolesterol biasana bodas, sareng batu anu dicampur coklat dina warna anu béda. Dina tahap awal formasi batu, asam chenodeoxycholic tiasa dianggo salaku ubar. Sakali dina hampru, éta laun-laun ngaleupaskeun batu koléstérol, tapi ieu prosés laun anu lumangsung sababaraha bulan. Atherosclerosis mangrupikeun patologi dicirikeun ku penampilan plé atherogenik dina permukaan jero témbok vaskular. Salah sahiji alesan utami pikeun ngembangkeun patologi sapertos kieu nyaéta ngalanggar kasaimbangan antara asupan koléstérol tina dahareun, sintésis sareng ékskrési tina awak. Pasien sareng atherosclerosis ngagaduhan dina konsentrasi LDL sareng VLDL. Aya hubungan tibalik antara konsentrasi HDL sareng kamungkinan ngembang atherosclerosis. Ieu konsisten sareng konsép fungsi LDL salaku operator koléstérol dina jaringan, sareng HDL tina jaringan. Dasar "prasyarat metabolik" pikeun pengembangan atherosclerosis nyaéta hiperkololémemia. (koléstérol luhur dina getih). Hypercholesterolemia berkembang: 1. Kusabab kaleuwihan asupan koléstérol, karbohidrat sareng lemak, 2. predisposisi genetik anu diwangun ku cacad turunan dina struktur LDL atanapi resépér apoB-100, kitu ogé dina ningkat sintésis atanapi sékrési apoB-100 (dina kasus hyperlipidemia gabungan kulawarga, dimana konsentrasi getih sareng koléstér sareng TAG dinaangkat luhur). Peranan penting dina mékanisme pangembangan atherosclerosis dimaénkeun ku modifikasi obat. Parobihan dina struktur lipid sareng protéin normal dina LDL ngajantenkeun aranjeunna ka luar nagri awak sahingga langkung diaksés ku néwak phagocytes. Modifikasi narkoba tiasa lumangsung ku sababaraha mékanisme: 1. glikosilasi protéin anu lumangsung nalika konsentrasi glukosa dina getih naék, 2. modifikasi péroxida, ngahasilkeun parobahan lipid dina lipoprotein sareng struktur apoB-100, 3. formasi kompléks otoimun tina LP-antibodi (ubar anu dirobih tiasa nyababkeun formasi autoantibodies). LDL anu dirobah diserep ku makrofag. Proses ieu henteu diatur ku jumlah diserep koléstérol, sapertos halna asupna kana sél ngalangkungan reséptor khusus, ku kituna makrofag dirobih sareng koléstérol sareng janten "sél foamy" anu tembus kana rohangan subendothelial. Hal ieu nyababkeun formasi bintik lipid atanapi dina lapisan témbok getih. Dina tahap ieu, endothelium vaskular tiasa ngajaga strukturna. Kalayan paningkatan jumlah busa busa, karusakan endothelial lumangsung. Karusakan nyumbang ka aktivasina platelet. Hasilna, aranjeunna ngaréduksi thromboxane, anu ngarangsang agrégasi trombosit, sareng ogé ngamimitian ngahasilkeun faktor pertumbuhan trombosit, anu ngarangsang proliferasi sél otot lemes. Kéngingna hijrah tina medial kana lapisan batin témbok artipérsial, sahingga nyumbang kana plak piagam. Lajeng piagam sprout sareng jaringan fibrous, sél dina mémbran fibrous aya necrotic, sareng koléstérol disimpen dina rohangan intercellular. Dina tahap pamekaran pamungkas, piagam teu kaeusi ku kalsium uyah sareng janten padet pisan. Di wewengkon plak, gumpalan getih sering wujud, meungpeuk lumen bejana, anu ngabalukarkeun gangguan sirkulasi akut dina situs jaringan anu pakait sareng perkembangan serangan jantung. Peraturan sintésis koléstérol - énzim konci (HMG-CoA reductase) dilakukeun ku sababaraha cara. Fosforilasi / dephosphorylasi HMG réduktase. Kalayan paningkatan dina rasio insulin / glukagon, fosforus template énzim ieu pas kana kaayaan anu aktip. Tumindak insulin dilaksanakeun ngalangkungan 2 énzim. HMG-CoA réduktase kinase fosfatase, anu kéngingkeun kinase janten kaayaan dephosforilasi anu henteu aktip: Fosfasease HMG-CoA réduktase ku ngarobih kana kaayaan aktif dephosphorylated. Hasil tina réaksi ieu mangrupikeun formasi bentuk aktif dephosforus HMG-CoA réduktase. Akibatna, dina nalika nyerep, koléstérér naék. Salila periode ieu, kasadiaan substrat awal pikeun sintésis koléstérol - acetyl - CoA nambahan (akibat tina tuangeun tuangeun tuangeun karbohidrat sareng lemak, saprak CoA acetyl kabentuk nalika ngarecah glukosa sareng asam lemak). Dina kaayaan postabsorbent, glukagon liwat proteingenase A ngrangsang fosforilasi HMG - CoA - réduktase, balik janten kaayaan anu teu aktip. Padamelan ieu ditingkatkeun ku kanyataan yén dina waktos anu sami, glukagon merangsang fosforilasi sareng teu aktivasi fosfatase HMG-CoA réduktase, ku kituna ngajaga HMG-CoA réduktase dina kaayaan aktif fosforil. Hasilna, sintésis koléstérol dina periode postabsorption sareng salami puasa dicegah ku sintésis endogen. Lamun eusi koléstérol dina dahareun dibawa ka 2%, maka sintésis kolégérogén endogenous menurun. Tapi gencatan sintésis koléstérol lengkep henteu lumangsung. Gelar ngahambat biosintésis kolésteris dina pangaruh koléstérol anu asalna tina dahareun bénten-béda ti jalma ka jalma. Ieu nunjukkeun personalitas tina prosés formasi koléstérol. Ku ngukur intensitas sintésis kolésterol, mungkin waé ngirangan konsentrasi dina getih. Lamun kasaimbangan antara asupan koléstérol sareng dahareun sareng sintésisna dina awak dina hiji sisi sareng ékskrési asam bili na koléstéris lain rusak, konsentrasi koléstérol dina jaringan sareng parobihan getih. Akibat anu paling serius pakait sareng paningkatan konsentrasi koléstérol getih (hypercholesterolemia), sedengkeun kamungkinan ngembangkeun aterosklerosis sareng kolelithiasis naek. Hiperkolesterolemia Familial (HCS) - formulir ieu langkung umum - sakitar 1 pasien per 200 urang. Cacat anu diwariskeun dina HCS nyaéta palanggaran nyerep LDL ku sél, sareng ku sabab panurunan dina katabolisme LDL. Salaku hasilna, konsentrasi LDL dina getih naék, ogé koléstérol saprak aya pisan di LDL. Kukituna, sareng HCS, déposisi koléstérol dina jaringan, hususna dina kulit (xanthomas), dina dinding arteri mangrupikeun ciri-ciri. Inhibition sintésis HMG-CoA réduktase Produk ahir jalur métabolik koléstérol. Éta ngurangan laju transkripsi gén HMG-CoA réduktase gén, sahingga ngahambat sintésisna nyalira. Ati aktip nyintésis asam bili tina koléstérol, sareng ku kituna asam bili ngahambat kagiatan gén Hédg-CoA. Kusabab HMG-CoA réduktase aya saatos sintésis sakitar 3, panyambutan sintésis énzim koléstérol ieu nyaéta pangaturan anu épéktip.
|