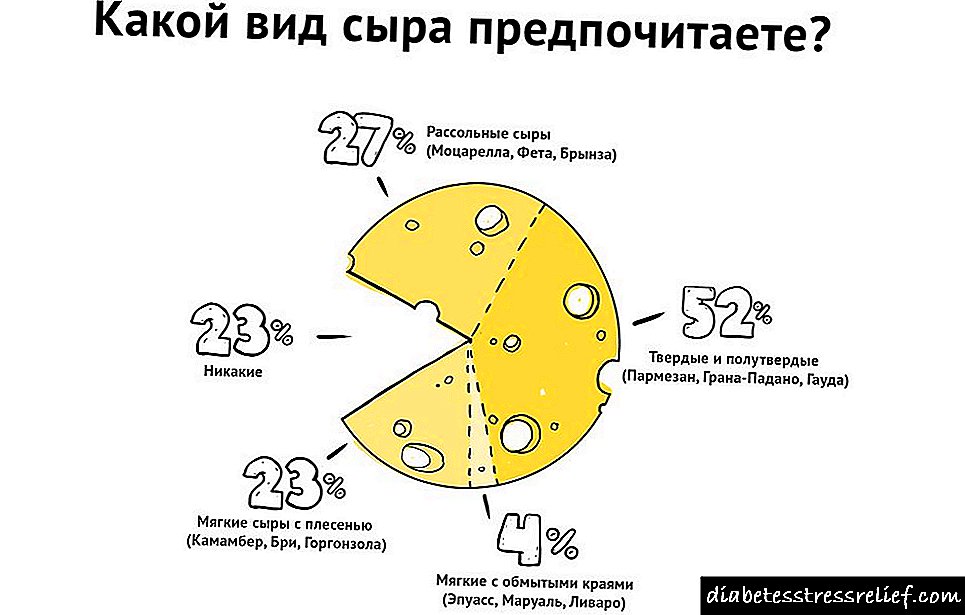
Biosintésis kolesterol sareng biokimia na - Diabetes
Tanpa ragu, koléstérol mangrupakeun lipid anu paling dipikanyaho pikeun masarakat umum; henteu kasohor sabab korélasi luhur antara kolesterol getih tinggi sareng frékuénsi panyakit kardiovaskular manusa. Kurang perhatian teu dibayar ka peran penting koléstérol salaku komponén mémbran sél sareng prékursor pikeun hormon stéroid sareng asam bili. Kolesterol dipikabutuh pikeun seueur sato, kalebet manusa, tapi ayana dina mamalia pangan tiasa pilihan - sél awak sorangan tiasa nyintésis tina prékursor saderhana.
Struktur sanyawa 27-karbon ieu nunjukkeun jalan kompléks pikeun biosintésisna, tapi kabéh atom karbonna disayogikeun ku prékursor tunggal - asetat. Blok Isoprene - perantaraan anu paling penting tina asétat ka koléstérol, aranjeunna prékursor seueur lipid alami, sareng mékanisme anu ngaleungitkeun blok isoprene sami dina sadaya jalur métabolik.
Kami dimimitian ku nguji tahap utama dina jalur biosintésis koléstéris ti asétat, teras bahaskeun transportasi koléstérol ngalangkungan aliran getih, diserep ku sél, pangaturan sintésis koléstéris normal, sareng régulasi dina kasus panyerepan atanapi pangangkut gangguan. Teras urang ningali bahan-bahan sanés anu asalna tina koléstérol, sapertos asam biliin sareng hormon stéroid. Tungtungna, pedaran ngeunaan jalur biosynthetic pikeun pembentukan sanyawa - turunan blok isoprene, dimana aya tahap awal umum kalayan sintésis kolésteris, bakal nunjukkeun kasipetan anu luar biasa tina kondensasi isoprenoid dina biosintésis.
Kolésterér ngahasilkeun tina acetyl-CoA dina opat tahap
Koléstérol, sapertos asam lemak ranté panjang, didamel tina acetyl-CoA, tapi pola pemasangan henteu béda pisan. Dina percobaan kahiji, asetét dilabélan ku 14 C boh dina atom métil atanapi karboksil ditambah kana pakan sato. Dumasar distribusi labél dina koléstérol terasing tina dua grup sato (Gbr. 21-32), hambalan énzimatik tina biosintésis kolésterol.
Gbr. 21-32. Sumber atom karbon koléstérol. Diidentipikasi nalika percobaan nganggo acétate radioaktif anu dilabélan nganggo métil karbon (hideung) atanapi karboksilat karbon (beureum). Dina struktur gondensasi, cincin dilambangkeun ku hurup A dugi D.

Sintésis lumangsung dina opat tahapan, sapertos dipidangkeun dina Gbr. 21-33: (1) résolusi tina tilu résidu asétat pikeun ngabentuk genep karbon panengah mevalonat, (2) konversi mevalonate pikeun diaktipkeun blok isoprene, (3) polimérisasi genep genep unit isoprén karbon pikeun ngabentuk squalene linear 30-karbon, (4) siklik kolonal pikeun ngabentuk opat cincin inti stéroid, dituturkeun ku parobihan parobihan (pangoksidasi, ngaleupaskeun atanapi migrasi gugus métil) kalayan pembentukan koléstérol.
Gbr. 21-33. Gambar umum biosintésis kolésteréris. Opat tahap sintésis dibahas dina téks. Blok Isoprene di squalene ditandaan ku garis luntur beureum.

Panggung (1). Sintésis mevalonat tina asetat. Tahap kahiji biosintésis kolesterol ngabalukarkeun pembentukan produk perantara mevalonate (Gbr. 21-34). Dua molekul Coetyl CoA condense to give acetoacetyl CoA, anu condenses with acetyl CoA molekul katilu pikeun ngabentuk sanyawa genep karbon β-hydroxy-β-methylglutaryl-CoA (HM G -CoA). Dua réaksi mimitina ieu dikatalisan thiolase sareng synthase NM G -CoA, masing-masing. Cytosolic Sintésis GM Co-CoA Jalur métabolik ieu béda ti isoenzyme mitokondria, anu ngatalisan sintésis NM G -CoA nalika kabentukna awak keton (tempo Gbr. 17-18).
Gbr. 21-34. Wangunan mevalonate ti acetyl-CoA. Sumber C-1 sareng C-2 mevalonate tina acetyl-CoA disorot dina warna pink.

Réaksi katilu watesan laju prosés. Di jerona, NM G -CoA diréduksi jadi mevalonate, anu masing-masing tina dua molekul NА D PH nyayogikeun dua éléktron. HMG-CoA réduksi protéin membran integral tina lancar ER, éta ngalayanan, sakumaha anu bakal urang tingali, salaku titik utama pangaturan jalur métabolik formasi koléstérol.
Panggung (2). Konvérsi mevalonate kana dua isoprén diaktipkeun. Dina tahap salajengna sintésis koléstérol, tilu gugus fosfat ditransfer tina molekul ATP jadi mevalonate (Gbr. 21-35). Fosfat kabeungkeut ka gugus hidroksil di C-3 mevalonate dina panengah 3-phospho-5-pyrophosphomevalonate nyaéta kelompok anu saé, dina lengkah saterusna kadua fosfat ieu sareng kelompok karboksilat anu padeukeut, ngabentuk beungkeut ganda dina produk lima-karbon ∆ 3 -isopentenyl pyrophosphate. Ieu mangrupikeun mimiti dua isoprén diaktipkeun - pamilon utama dina sintésis koléstérol. Isomerisasi Δ 3 -isopentenylpyrophosphate masihan isoprene anu diaktipkeun kadua dimethylallyl pyrophosphate. Sintésis isopentenyl pyrophosphate dina sitoplasma sél tutuwuhan lumangsung dumasar kana jalur anu dijelaskeun di dieu. Tapi, kloroplas tutuwuhan sareng seueur baktéri nganggo jalur bebas tina mevalonate. Rute alternatif ieu henteu kapendakan dina sato, janten pikaresepeun nalika nyiptakeun antibiotik anyar.
Gbr. 21-35. Konversi mevalonate kana blok isoprén diaktipkeun. Genep unit diaktipkeun ngagabungkeun pikeun ngabentuk squalene (tempo Gambar 21-36). Grup ninggalkeun 3-phospho-5-pyrophosphomevalonate disorot dina warna pink. Dina kurung alun-alun mangrupikeun perantara hipotesis.

Panggung (3). Kondensasi genep unit isoprén diaktipkeun pikeun ngabentuk squalene. Isopentenyl pyrophosphate sareng dimethylallyl pyrophosphate ayeuna ngalaman kondensasi sirah-to-buntut, dimana hiji gugus pyrophosfat ngalir sareng bentuk ranté 10-karbon - geranyl pyrophosphate (Gbr. 21-36). (Pyrophosphate nempel kana sirah.) Geranyl pyrophosphate ngalaman kondensasi sirah-to-buntut di handap ieu sareng isopentenyl pyrophosphate, sareng bentuk panengah 15-karbon farnesyl pyrophosphate. Ahirna, dua molekul farnesyl pyrophosphate ngagabung "sirah ka sirah", duanana gugus fosfat dicabut - kabentuk squalene
Gbr. 21-36. Formasi Squalene. Struktur squalene anu ngandung 30 atom karbon lumangsung nalika kondensasi berturut-turut diaktipkeun ku blok isoprén (lima-karbon).

Ngaran umum pikeun perantara ieu asalna tina ngaran sumber ti mimiti diasingkeun. Geraniol, salah sahiji komponén minyak mawar, ngagaduhan rasa geranium, sareng farnesol, kapanggih dina warna akasia farnesa, ngagaduhan sanés tina aroma lebak. Seueur bau tutuwuhan alami kanggo sanyawa diwangun tina blok isoprén. Squalene, mimiti diasingkeun tina ati hiu (Spésiés Squalus), diwangun ku 30 atom karbon: 24 atom dina ranté utama sareng genep atom dina meungkeut logam.
Panggung (4). Transformasi squalene jadi opat cincin tina inti stéroid. Dina ara. 21-37 jelas katingal yén struktur rantai squalene, sareng sterol - siklik. Sakabéh sterol gaduh opat cincin anu condensasi anu ngabentuk inti stéroid, sareng sadaya alkohol aya gugus hidroksil dina atom C-3, ku kituna nami inggris Inggris. Dina kaayaan tindakan ménalxygenase squalene hiji atom oksigén ti O ditambah kana tungtung rantay squalene 2 sareng hiji epoxide kabentuk. Énzim ieu mangrupikeun oksidase campuran-fungsi sanésna (tambihan 21-1), NADPH ngurangan atom oksigén séjén tina O 2 pikeun H2 O. Produk Ganda Deui squalene-2,3-epoxide diatur supados réaksi anu lumayan konsisten tiasa kéngingkeun ranté epoxide squalén janten struktur siklik. Dina sél sato, siklusisasi ieu nuju lanosterol anu ngandung opat cincin ciri inti inti. Hasilna, lanosterol dirobih kana koléstérol ngalangkungan 20 réaksi salebetna réaksi, anu kalebet migrasi sawaréh gugus logam sareng ngaleupaskeun batur. Katerangan ngeunaan biosintésis anu luar biasa ieu, salah sahiji anu paling hese diantarana, dipasihkeun ku Conrad Bloch, Theodore Linen, John Cornfort sareng George Popiak dina akhir taun 1950-an.
Gbr. 21-37. Panutupan cincin janten liniér liniér janten inti stéstor. Tahap kahiji katalis ku oksidase nganggo fungsi dicampur (muslanxygenase), anu kosubstrat nyaéta N AD PH. Produkna mangrupikeun epoksida, anu dina tahap salajengna janten siklus pikeun ngabentuk inti stéroid. Produk ahir réaksi ieu dina sél sato nyaéta koléstérol; dina organisme séjén rada béda-béda dibentukna.


Kolesterol mangrupakeun ciri sterol sél sato, tutuwuhan, jamur sareng protés ngahasilkeun stérol anu sami pisan.
Aranjeunna nganggo rute sintésis anu sami kana squalene-2,3-epoxide, tapi teras jalurna rada saeutik, sareng stona sanésna kabentuk, sapertos sigmasterol dina seueur pepelakan sareng ergosterol dina fungi (Gbr. 21-37).
Conto Biaya Énergi 21-1 pikeun Sintésis Squalene
Naon biaya énérgi (dinyatakeun minangka ATP molekul) pikeun sintésis hiji molekul squalene?
Solusi. Dina sintésis squalene tina acetyl-CoA, ATP ukur ukur di panggung nalika mevalonate dirobah jadi prékursor isaléna anu diaktipkeun. Genep molekul isoprén anu diaktipkeun diperyogikeun pikeun ngawangun molekul squalene, sareng tilu molekul ATP diwajibkeun ngahasilkeun unggal molekul diaktipkeun. Dina total, 18 molekul ATP dikaluarkeun dina sintésis hiji molekul squalene.
Sanyawa koléstérol dina awak
Dina vertebrata, jumlah koléstérol ageung disintésis dina ati. Sababaraha kolésteris disintésis aya dilebetkeun kana mémbran hépatosit, tapi biasana diékspor dina salah sahiji tina tilu bentuk: kolésterilium (bili), asam biliin atanapi éster koléstérol. Asam bili sarta uyah nyaéta turunan hidrofilik tina koléstérol, anu disintésis dina ati sareng nyumbang kana nyerna lipid (tingali Gbr. 17-1). Éster koléstérol ngawujud dina haté ku polah acil-CoA-koléstér-acyltransferase (ACAT). Énzim ieu ngatalisan transisi sésana asam lemak tina koénzim A ka gugus hidroksil (Gbr. 21-38), janten koléstérol janten bentuk langkung hidrofobik. Éster kolesterol dina partikel lipoprotein anu disékrésikeun diangkut ka jaringan sanés nganggo koléstérol atanapi disimpen dina ati.
Gbr. 21-38. Sintésis éster koléstérol Éterifikasi ngajadikeun koléstérol minangka bentuk hidrofobik langkung pikeun neundeun sareng transportasi.

Kolesterol dipikabutuh pikeun sadaya jaringan organisme sato ngembang pikeun sintésis mémbran, sareng sababaraha organ (contona, kelenjar adrenal sareng kelenjar kelamin) nganggo koléstérol salaku prékursor pikeun hormon stéroid (ieu bakal dibahas di handap). Kolesterol ogé prékursor pikeun vitamin D (tingali Gambar 10-20, v. 1).
Kolesterol sareng lipid séjén mawa lipoproteins plasma
Éster kolesterol sareng koléstérol, sapertos triacylglycerol sareng fosfolipid, sacara praktis teu larut dina cai, kumaha, aranjeunna kedah ngalih tina jaringan dimana aranjeunna disintésis kana jaringan dimana éta bakal disimpen atanapi dikonsumsi. Éta anu dibawa ku aliran getih dina bentuk lipoproteins plasma getih - kompleks macromolecular protéin pamawa khusus (apolipoproteins) kalayan fosfolipid, koléstérol, éster koléstérol sareng triacylglycerol anu aya dina kompleks ieu dina sababaraha kombinasi.
Apolipoproteins ("apo" ngarujuk kana protéin bebas lipid sorangan) ngagabung sareng lipid pikeun ngabentuk sababaraha fraksi partikel lipoprotein - kompleks spherical ku lipid hidrofobik di tengah sareng ranté asam amino hydrophilic dina permukaan (Gbr. 21-39, a). Kalayan sagala rupa kombinasi lipid sareng protéin, partikel tina kapadetan anu béda-béda dibentuk - tina chylomicrons ka lipoproteins dénsitas tinggi. Partikel ieu tiasa dipisahkeun ku ultracentrifugation (Tabél 21-1) sareng dititénan sacara visual nganggo mikroskop éléktron (Gambar 21-39, b). Unggal fraksi lipoprotein ngalaksanakeun fungsi husus, anu ditangtukeun ku tempat sintésis, komposisi lipid sareng eusi apolipoprotein. Sahenteuna 10 béda apolipoprotein kapanggih dina plasma getih manusa (Tabél 21-2), anu beda-beda dina ukuran, réaksi ku antibodi khusus, sareng distribusi karakteristik dina sababaraha kelas lipoprotein. Komponén protéin ieu mangrupikeun bahan sinyal pikeun ngarahkeun lipoprotein ka jaringan spésifik atanapi ngaktifkeun énzim anu meta dina lipoproteins.
Méja 21-1. Lipoproteins plasma manusa
Komposisi (fraksi massa,%)
r = 513,000). Zat LDL ngandung inti kira-kira 1,500 molekul éster koléstérol, di sabudeureun inti aya cangkang 500 molekul koléstérol, 800 molekul fosfolipid sareng hiji molekul apoB-100. b - opat kelas lipoprotein, katingali ku mikroskop éléktron (saatos wujud anu négatip). Jarum jam, dimimitian ti sosok kénca luhur: chylomicrons - kalayan diaméter 50 dugi 200 nm, PL O NP - ti 28 dugi 70 nm, HDL - ti 8 dugi 11 nm, sareng LDL - ti 20 dugi ka 55 nm. Sipat lipoprotein dirumuskeun dina tabél. 21-2.

Chylomicrons, disebut dina Sec. 17, mindahkeun kadaharan triacylglycerol tina peujit ka jaringan sanés. Ieu mangrupikeun lipoprotein anu panggedéna, aranjeunna gaduh kapadetanna panghandapna sareng eusi relatif paling luhur tina triacylglycerol (tempo Gbr. 17-2). Chylomicrons disintésis dina er sél épitélial anu ngajamin peujit leutik, teras ngalirkeun sistem limfatik sareng asupkeun aliran getih ngaliwatan urat subclavian kénca. Apolipoprotein Chylomicron ngandung apoB-48 (unik pikeun kelas lipoproteins ieu), apoE sareng apoC-II (Tabél 21-2). AroC-II ngaktifkeun lipoprotein lipase dina kapiler jaringan adipose, jantung, otot rangka sareng laktat kelenjar susu, mastikeun aliran asam lemak bébas kana jaringan ieu. Ku kituna, chylomicrons mindahkeun asam lemak dahareun ka jaringan, dimana éta bakal dikonsumsi atanapi disimpen sapertos suluh (Gbr. 21-40). Résidu Chylomicron (utamana dibébaskeun tina triacylglycerol, tapi tetep ngandung koléstérol, apoE sareng apoB-48) diangkut ku aliran getih kana ati. Dina ati, reséptor ngabeungkeut apoE anu aya dina résidu chylomicron sareng nyéépkeun panyerepan ku endocytosis. Dina hépatosit, résidu ieu ngaleupaskeun koléstérol anu dikandung sareng dirusak di lisina.
Méja 21-2. Manusa plasma lipoprotein apolipoproteins
Fungsi (upami dikenal)
Ngaktipkeun L CAT, hubungan sareng transporter ABC
Nunjukkeun L CAT
Ngaktipkeun L CAT, transportasi koléstérol / clearance
Ngarasakeun ka reséptor LDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Ngajalankeun pelepasan VLDL sareng résidu chylomicron
Nalika dahareun ngandung langkung asam lemak ti ayeuna tiasa dianggo salaku bahan bakar, aranjeunna janten triacylglycerol dina ati, anu ngawangun fraksi sareng apolipoproteins khusus lipoproteins dénsitas pisan low (VLDL). Karbohidrat kaleuleuwihan dina ati ogé tiasa dirobih kana triacylglycerol sareng diékspor salaku VLDL (Gbr. 21-40, a).Salian triacylglycerol, fraksi VLDL ngandung jumlah éster kolésterol sareng koléstérol, ogé apoB-100, apoC-1, apoC-II, apoC III sareng apoE (Tabél 21-2). Lipoprotein ieu ogé diangkut ku getih tina ati ka otot sareng jaringan adipose, dimana, saatos lipoprotein lipase diaktipkeun ku apo-C II, asam lemak bébas dileupaskeun tina triacylglycerol tina fraksi VLDL. Adipocytes néwak asam lemak bébas, teras janten deui kana triacylglycerol, anu disimpen dina sél ieu dina bentuk inpormasi lipid (tetes), mios, sabalikna, langsung ngoksidasi asam lemak pikeun ngahasilkeun énergi. Kaseueuran résidu VLDL dikaluarkeun tina sirkulasi ku hépatosit. Nyerepna, sami sareng nyerep chylomicrons, ditengah ku reséptor sareng gumantung ayana apoE di résidu VLDL (tambihan. 21-2, hubungan antara apoE sareng Panyakit Alzheimer dijelaskeun).
Gbr. 21-40. Lipoproteins sareng transportasi lipid, sareng - lipid diangkut ku aliran getih dina bentuk lipoprotein, anu digabungkeun kana sababaraha fraksi anu gaduh fungsi anu béda sareng komposisi anu béda sareng protéin sareng lipid (tab. 21-1, 21-2) sareng pakait kana kapadetan fraksi ieu. Lipid dahareun dikumpulkeun kana chylomicrons, kalolobaan triacylglycerol anu aya di antarana dikaluarkeun ku lipoprotein lipase kana adipose sareng jaringan otot dina kapilér. Sésa Chylomicron (ngandung protéin sareng koléstérol) kawengku ku hépatosit. Lipid endogenous sareng koléstérol na ati dikirimkeun ka jaringan adipose sareng otot dina bentuk VLDL. Pelepasan lipid ti VLDL (bareng sareng leungitna sababaraha apolipoproteins) laun-laun ngarobah VLDLP ka LDL, anu ngirim koléstérol kana jaringan extrahepatic atanapi mulih deui kana ati. Ati ngarawat sésa-sésa VLDL, LDL sareng sésa-sésa chylomicrons ku endocytosis mediator-medi. Kaleuwihan koléstérol dina jaringan extrahepatic diangkut deui kana ati dina bentuk LDL. Dina ati, bagian koléstérol robah jadi uyah bili. b - conto plasma getih anu dicandak saatosna kalaparan (kénca) sareng saatos tuangeun tuangeun ku eusi gajih (katuhu katuhu). Chylomicrons dibentuk ku tuang tuangeun lemak masihan plasma salaku kasaruaan éksternal susu.

Kalayan leungitna triacylglycerol, bagian tina VLDL dirobih kana résidu VLDL, anu ogé disebut lipoproteins perantara pertengahan (VLDL), ngaleungitkeun salajengna triacylglycerols ti VLDL masihan lipoproteins dénsitas low (LDL) (tab. 21-1). Fraksi LDL, anu beunghar pisan dina éster koléstérol sareng koléstérol, sareng ogé ngandung apoB-100, ngirimkeun koléstérol dina jaringan extrahepatic anu nyandak reséptor khusus sadar apoB-100 dina mémbran plasma. Reséptor ieu nyapihkeun karangan éster koléstérol sareng éster (sapertos anu dijelaskeun di handap).
Tambahan 21-2.Alél ApoE nangtoskeun kajadian tina kasakit Alzheimer
Dina populasi manusa, aya tilu varian anu dipikanyaho (tilu alél) tina génod anu ngeposip apolipoprotein E. Tina alél apo-laré, alél APOEZ paling umum dina manusa (sakitar 78%), alél APOE4 sareng APOE2 masing-masing masing-masing 15 sareng 7%. Allele APOE4 hususna ciri jalma anu boga panyakit Alzheimer, sareng hubungan ieu ngamungkinkeun ngaduga lumangsungna panyakit kalayan kamungkinan anu luhur. Jalma anu ngawariskeun APOE4 ngagaduhan résiko ageung kaserang panyawat Alzheimer. Jalma homozygous pikeun APOE4 aya 16 kali langkung dipikaresep ngembangkeun panyakit, umur rata-rata jalma anu sakitar umurna sakitar 70 taun. Pikeun jalma anu ngawariskeun dua salinan AROEZ, sabalikna, umur rata-rata panyawat Alzheimer ngaleuwihan 90 taun.
Dasar molekularna pakaitna antara apoE4 sareng panyawat Alzheimer tacan kanyahoan. Salaku tambahan, éta masih henteu jelas kumaha apoE4 tiasa mangaruhan tumuh tali amyloid, anu tétéla panyababna panyawat Alzheimer (tingali Gbr. 4-31, v. 1). Asumsi fokus dina peran apoE dina kamungkinan stabilisasi struktur sitoskeleton neuron. Protokol apoE2 sareng apoEZ meungkeut sajumlah protéin anu terkait sareng mikrotubul saraf, sedengkeun apoE4 henteu ngabeungkeut. Ieu bisa ngagancangkeun maotna neuron. Naon waé yén mékanisme ieu, pangamatan ieu méré harepan pikeun ngembangkeun pamahaman kami ngeunaan fungsi biologis apolipoproteins.
Jenis kaopat lipoprotein - lipoproteins dénsitas tinggi (HDL), fraksi ieu dibentuk dina ati sareng peujit leutik dina bentuk partikel anu beunghar protéin anu ngandung saeutik koléstérol sareng bébas éster koléstér (Gbr. 21-40). Fraksi HDL ngandung apoA-I, apoC-I, apoC-II sareng apolipoproteins sanés (Tabél 21-2), kitu ogé lecithin-kolesterol-acyltransferase (LC AT), anu ngatalisan kabentukna éster koléstérin tina lésitin (fosfatidilkol) sareng koléstérol (Gbr. 21-41). L CAT dina beungeut partikel HDL anu nembé dibentuk ngarobah koléstor chylomicron sareng fosfatidilchol sareng VLDL jadi éster koléstérol, anu mimiti ngabentuk inti, ngarobih partikel HDL discoid anu nembé kabentuk janten partikel HDL spherical asak. Lipoprotein anu beunghar koléstérol ieu teras balik deui ka ati, dimana koléstérol "dikaluarkeun", sababaraha koléstérol ieu dirobih jadi uyah bili.
Gbr. 21-41. Réaksi dikandung ku lecithin-kolesterol-acyltransferase (L CAT). Énzim ieu aya dina permukaan partikel HDL sareng diaktipkeun ku apoA-1 (komponén tina fraksi HDL). Éster kolesterol ngumpulkeun di jero partikel HDL anu nembé kabentuk, janten kana HDL dewasa.

HDL tiasa diserep dina ati ku endocytosis diturunkeun ku reséptor, tapi sahenteuna sababaraha koléstérol HDL dikirim ka jaringan sanés ku mékanisme anu sanés. Partikel HDL tiasa ngabeungkeut protéin SR - BI dina mémbran plasma sél ati sareng dina jaringan stéroid sapertos kelenjar adrénal. Reséptor ieu henteu mediasi endocytosis, tapi transfer partial sareng selektif koléstérol sareng lipid HDL sanésna kana sél. Fraksi "dikonsumsi" HDL teras deui asup aliran getih, dimana éta ngalebetkeun bagian anyar lipid ti chylomicrons sareng résidu VLDL. HDL anu sami ogé tiasa nyandak koléstérol anu disimpen dina jaringan extrahepatic sareng mindahkeun kana ati ku ngabalikeun angkutan koléstérol (Gbr. 21-40). Dina salah sahiji varian angkutan sabalikna, interaksi HDL anu ngagaduhan hasilna sareng reséptor SR-BI dina sél anu beunghar kolésterol ngamimitian penyebaran pasip tina permukaan sél kana partikel HDL, anu teras mindahkeun koléstérol ka ati. Dina varian angkutan sabalikna deui dina sél koléstérol anu saé, saatos dipisahkeun HDL, apoA-I interaksi sareng transporter aktif, protein ABC. ApoA-I (sareng sigana HDL) kaserep ku endokosisosis, teras disumputkeun deui, sarat ku koléstérol, anu diangkut kana ati.
Protein ABC1 mangrupikeun bagian tina kulawarga anu ageung seueur operator, sabab operator éta kadangkala disebut transportasi ABC, sabab sadayana ngandung kasét-iketan ngiket ATP (ATP - kaset ngariung), aranjeunna ogé gaduh dua domain transmembrane kalayan genep helem transmembrane (tingali bab. . 11, v. 1). Protéin ieu aktip mindahkeun seueur ion, asam amino, vitamin, hormon stéroid sareng uyah bili ngaliwatan mémbran plasma. Perwakilan pamawa kulawarga sanésna nyaéta protein CFTR, anu, ku fibrosis sista, ruksak (tingali tambihan 11-3, v. 1).
Éster kolesterol asup kana sél ngalangkungan endocytosis dianté ku reséptor
Masing partikel LDL dina aliran getih ngandung apoB-100, anu diakui ku protéin reséptor permukaan anu khusus -Reséptor LDL dina mémbran sél anu peryogi néwak koléstérol. Beungkeutan LDL ka reséptor LDL ngamimitian endokytosis, sabab LDL sareng reséptor na pindah kana sél dina jero endosom (Gbr. 21-42). Pungkasan endosome ngiringan sareng lysosome, anu ngandung énzim anu nghidrolisis éster koléstérol, ngaluarkeun koléstérol sareng asam lemak kana sitosol. ApoB-100 ti LDL ogé ngarobih pikeun ngabentuk asam amino anu dirembes kana sitosol, tapi reséptor LDL nyingkahan degradasi sareng wangsul deui kana permukaan sél pikeun ilubiung deui dina perbuatan LDL. ApoB-100 ogé aya di VLDL, tapi domain anu mengikat reseptor henteu tiasa ngabeungkeut ku reséptor LDL; konvérsi VLDLP ka LDL ngajantenkeun domain reséptor anu mengikat kana apoB-100 diaksés. Jalur angkutan kolesterol getih ieu sareng endokosisosis édisium reséptor na dina jaringan target anu ditaliti ku Michael Brown sareng Joseph Goldstein.
Michael Brown sareng Joseph Goldstein

Gbr. 21-42. Nangkep koléstérol ku endocytosis diturunkeun reséptor.

Kolesterol, anu asup kana sél ku cara kieu, tiasa diasupkeun kana mémbran atanapi didéplérasi deui ku ACAT (Gbr. 21-38) kanggo disimpen dina sitosol jero ogé lipid lipid. Nalika aya cukup koléstérol anu aya dina fraksi LDL getih, akumulasi koléstérol intrélular kaleuleuwihan dicegah ku ngirangan laju sintésisna.
Reséptor LDL ogé ngabeungkeut apoE sareng gaduh peran anu penting dina ngalaksanakeun chylomicrons sareng résidu VLDL ku ati. Nanging, upami reséptor LDL henteu sayogi (sapertos, dina galur beurit sareng gén reséptor LDL anu leungit), résidu VLDL sareng chylomicrons masih kaserep ku ati, sanaos LDL henteu nyerep. Ieu nunjukkeun ayana sistem cadangan bantu pikeun endokosisosis édisium ditengah antara VLDL sareng résidu chylomicron. Salah sahiji resép anu cagar nyaéta protéin LRP (protéin nu aya hubunganana sareng lipoprotein), anu aya hubunganana sareng reséptor lipoprotein, anu ngiket kana apoE sareng sajumlah ligan sanésna.
Sababaraha tingkat régulasi biosintésis kolésterés
Sintésis kolesterol mangrupikeun prosés anu rumit sareng énérgi, ku kituna jelas yén awak aya mangpaat pikeun boga mékanisme pikeun ngatur biosintésis kolésterés, anu nambihan deui sajumlah naon anu ngahasilkeun dahareun. Dina mamalia, produksi koléstér diatur ku konsentrasi intrasélular
kolesterol sareng hormon glukagon sareng insulin. Tahap konversi HMG - CoA ka mevalonate (Gbr. 21-34) ngawates laju dina jalur métabolik formasi koléstérol (titik utama régulasi). Réaksi ieu dikandung ku HMG - CoA réduktase. Peraturan minangka réspon kana parobahan koléstérol ditengah ku sistem pangaturan transkrip anu elegan pikeun sérmode génoding HMG - CoA. Gen ieu, sareng langkung ti 20 énzim énzim sanés anu aub dina nyerep sareng sintésis koléstérol sareng asam lemak tak jenuh, dikawasa ku kulawarga leutik protéin anu disebut protéin anu berinteraksi sareng unsur sterol-pangaturan pembentukan protéin (SREBP, unsur pangaturan unsur sterol) . Saatos sintésis, protéin ieu diwanohkeun ka reticulum endoplasmic. Hiji-hijina fungsi domain SREBP amino-leyur salaku aktivator transkripsi nganggo mékanisme anu dijelaskeun dina Ch. 28 (v. 3). Nanging, domain ieu henteu ngagaduhan aksés kana inti sareng henteu tiasa ngiringan aktivasina gén salami tetep dina molekul SREBP. Pikeun ngaktipkeun transkripsi gén HMG - CoA réduktase sareng génér anu sanés, domain anu aktif transkrippis dipisahkeun tina sesa SREBP ku cleavage proteolitik. Nalika koléstérol tinggi, protéin SREBP henteu aktif, tetep dina ER di kompleks sareng protéin anu sanésna disebut SCAP (SREBP - protéksi ngilangkeun protéin) (Gbr. 21-43). Éta SCAP anu ngabeungkeut koléstérol sareng sajumlah sterol sanésna, anu bertindak minangka sensor stérol. Nalika tingkat tina sterol tinggi, kompleks SCAP - SREBP sigana hubungan sareng protéin anu sanés, anu ngajaga sakabeh kompléks dina ER. Nalika tingkat tina suhu dina sél turun, parobahan konformasional di SCAP nyababkeun leungitna aktipitas, sareng kompléks SCAP - SREBP hijrah di jero vesikel ka kompleks Golgi. Dina kompleks Golgi, protéin SREBP dibuuhan dua kali ku dua protease béda-béda, cleavage kadua ngaleupaskeun domain amino-terminal kana sitosol. Domain ieu gerak ka inti sareng ngaktifkeun transkripsi gén target. Domain protein-SREBP amino-terminal ngagaduhan satengah hirup anu pondok sareng gancang diturunkeun ku proteasomes (tempo Gbr. 27-48, t. 3). Nalika tingkat sterol naékna cekap, sékrési proteolitik tina protéin protéin EBP SR sareng terminus amino dipeungpeuk deui, sareng degradasi proteasome domain aktif anu aya ngajantenkeun gancangna gen sasaran.
Gbr. 21-43. Aktipkeun of SR EBP. Protéér SREB P ngainteraksi sareng unsur anu ngatur régolasi (warna héjo), langsung saatos sintésis, diwanohkeun ka ER, ngabentuk kompleks sareng S CAP (warna beureum). (N jeung C nunjukkeun harga amina sareng karboksil dina protéin.) Dina kaayaan S-CAP, protéin SRE BP henteu aktif. Nalika tingkat sterol turun, kompleks SR EBP-S CAP hijrah ka komplek Golgi, sareng protéin SR EBP sacara berturut-turut dibentuk ku dua protease anu béda-béda. Domain protéin bébas anu dibebaskeun dina protéin protéin EBP migrasi ka inti, dimana ngaktifkeun transkripsi gén sterol-diatur.

Sintésis kolesterol ogé diatur ku sababaraha mékanisme séjén (Gbr. 21-44). Kontrol hormonal ditengah ku modifikasi kovalén tina réduktase NM G-CoA. Énzim ieu aya dina bentuk fosforil (teu aktif) sareng formosfosorasi (aktip). Glucagon ngrangsang fosforilasi (inactivation) énzim, sareng insulin promosi dephosphorylation, ngaktifkeun énzim sareng milih sintésis koléstérol. Konsentrasi kolésterol intrasélular anu luhur ngaktifkeun ASAT, anu ningkatkeun ésterisasi koléstérol pikeun déposisi. Tungtungna, tingkat koléstérol sélulér anu luhur ngahambat transkripsi gén anu nyerep hiji reséptor LDL, ngirangan produksi reséptor ieu sareng, saba kitu, karangan koléstérol tina getih.
Gbr. 21-44. Perda kadar kolésterol kasaimbangan antara sintésis sareng nyerep koléstérol ti dahareun. Glucagon mempermudah fosforilasi (inactivation) tina NM G -CoA réduktase, insulin promosi dephosphorylation (aktivasi). Métabolit kolesterol X anu teu dikenal anu merangsang proteolisis tina NM G -CoA réduktase.

Kolesterol teu teratur bisa ngakibatkeun panyawat serius di manusa. Nalika jumlah kolésterol sareng koléstésis total tina réstoran diperyogikeun ngalegaan jumlah anu diperyogikeun pikeun Déwan membran, sintésis uyah bili na stéroid, akumulasi patologis koléstér dina pembuluh darah (plak atherosclerotic) bisa muncul, ngarah kana sumbatan (atherosclerosis). Di nagara-nagara industrialisasi, éta kagagalan jantung kusabab halangan tina arteri koronér anu jadi panyabab utama kana pati. Perkembangan atherosclerosis aya hubunganana sareng kadar kolesterol tinggi darah, khususna sareng koléstérol tinggi ditolerir ku fraksi LDL; tingkat luhur getih HDL, sabalikna, nguntungkeun mangaruhan kaayaan pembuluh darah.
Kalayan hypercholesterolemia turunan (cacat genetik), tingkat koléstérol getih pisan tinggi - atherosclerosis parna parah di jalma-jalma ieu anu aya di budak leutik. Kusabab tina reséptor LDL anu cacat, panerapan édisién-éntuk anu ditampi dina kolesterol LDL lumangsung. Hasilna, koléstérol henteu dikaluarkeun tina aliran getih, éta éta akumulasi sareng nyumbang kana formasi plak atherosclerotic. Sintésis koléstor endogen terus, najan kaleuwihan koléstérol dina getih, sabab koléstrasélular teu tiasa lebet kana sél pikeun ngatur sintésis intrasélular (Gbr. 21 -44).Pikeun pengobatan pasien kalayan hypercholesterolemia turunan sareng panyakit sanés anu aya hubunganana sareng koléstum sérum anu luhur, kelas statin dianggo. Sababaraha di antarana dicandak tina sumber alami, sedengkeun anu sanés disintésis ku industri farmasi. Statins sami sareng mevalonate (tambihan 21-3) sareng sambetan kalapa tina NMS-CoA reductase.
Addendum 21-3. PERÉKARA. Hipotesis lipid sareng nyiptakeun statin
Panyakit jantung koronér (CHD) mangrupikeun panyebab utama mortir di nagara-nagara maju. Penyempitan arteri koronér anu ngalaksanakeun getih ka jantung balukar tina pembentukan deposit lemak anu disebut plak atherosclerotic; plak ieu ngandung koléstérol, protéin fibrillar, kalsium, gumpalan platelet, sareng sempalan sél. Dina abad XX. Aya debat aktif ngeunaan hubungan antara halangan arteri (atherosclerosis) sareng kolesterol getih. Diskusi ieu sareng panalungtikan aktip dina arah ieu parantos nyiptakeun ubar mujarab anu nurunkeun koléstérol.
Dina taun 1913, N.N. Anichkov, saurang ilmuwan Rusia anu terkenal sareng ahli dina patologi ékspérimén, medarkeun hiji karya dimana anjeunna nunjukkeun yén arnab anu disuurkeun ku tuang beunghar kolesterol ngembangkeun karusakan kana pembuluh darah anu nyarupaan plak atherosclerotic dina kapal-kapal sepuh. Anichkov ngalaksanakeun panalitianana salami sababaraha taun sareng nyebarkeun hasil dina jurnal Kulon anu kaceluk. Hanjakalna, data na henteu janten dasar modél pangwangunan atherosclerosis di manusa, saprak dina waktos éta hipotesis aya yén panyakit ieu mangrupikeun hasil alami tina sepuh sareng teu tiasa dicegah. Nanging, bukti laun-laun ngumpulkeun hiji hubungan antara koléstum sérum sareng kamekaran atherosclerosis (hipotésis lipid), sareng taun 1960-an. sababaraha peneliti sacara eksplisit nyatakeun yén panyakit ieu tiasa diubaran ku pangobatan. Nanging, sudut pandang tibalik aya dugi dugi ka publikasi dina taun 1984 tina hasil panilitian anu lega ngeunaan peran koléstérol anu dilakukeun ku Amérika National Institute of Health (Trialary Primary Pencegahan Pencegahan). Panurunan signifikan statistik dina frékuénsi infark miokardial sareng stroke kalayan panurunan dina kolesterol getih nunjukkeun. Dina ulikan ieu, koléstérol, résin bursa anion anu ngabeungkeut asam bili, dipaké pikeun nurunkeun koléstérol. Hasilna geus ngrangsang milarian obat terapi anyar anu langkung kuat. Kuring kedah nyarios yén di dunya ilmiah, mamang ngeunaan validitas hipotésis lipid ngaleungit ngan ukur kalayan munculna statin dina ahir taun 1980an - awal taun 1990an.
The statin mimiti kapanggih ku Akira Endo di Sankyo di Tokyo. Endo nyebarkeun karyana dina 1976, sanaos anjeunna ngatasi masalah métabolisme koléstérér salami sababaraha taun. Dina taun 1971, anjeunna nyarankeun yén sambetan sintésis koléstérol ogé dikandung dina produsén supa antibiotik anu diulik dina waktos éta. Pikeun sababaraha taun damel sacara intensif, anjeunna nganalisis langkung ti 6,000 budaya ti sagala rupa suung, dugi ka anjeunna hasil anu positif. Sanyawa anu dihasilkeun disebut compactin. Zat ieu ngagaduhan koléstérol anjing sareng monyét. Panaliti ieu narik perhatian Michael Brown sareng Joseph Goldstein Universitas Sakola Médis Texas Southwestern. Brown sareng Goldstein, sareng sareng Endo, ngamimitian diajar babarengan sareng mastikeun data na. Kasuksésan utama uji klinis munggaran aub kalebet perusahaan farmasi dina ngembangkeun narkoba anyar ieu. Di Merck, tim anu dipimpin ku Alfred Alberts sareng Roy Wagelos ngaluncurkeun saringan anyar kabudayaan supa sareng, salaku hasil tina nganalisa 18 budaya, mendakan ubar aktip anu sanés. Zat anu anyar disebut lovastatin. Sanajan kitu, dina waktos anu sami, seueur dipercaya yén administrasi dosis tinggi compactin ka anjing nyababkeun ngembangkeun kanker sareng milarian statin énggal dina taun 1980-an. parantos ditunda. Nanging, ku waktos éta, pedah ngagunakeun statin pikeun ngubaran pasien sareng hypercholesterolemia familial parantos jelas. Saatos seueur konsultasi sareng ahli internasional sareng Administrasi Pangan sarta Narkoba (FDA, AS), Merck mimiti ngembangkeun lovastatin. Studi anu éksténsif salami dua puluh taun ka hareup parantos ngungkabkeun pangaruh karsinogenik sareng lovastatin sareng ubar énggal anu anyar muncul.
Gbr. 1. Statins mangrupakeun sambetan NM G-CoA réduktase. Perbandingan struktur mevalonat sareng opat produk farmasi (statin) anu ngahambat tindakan NM G -CoA réduktase.

Statins ngahambat tina tindakan HMG - CoA - réduktase, nyontoan struktur mevalonate, sareng ku kituna ngahalang sintésis koléstérol. Dina pasien anu hypercholesterolemia disababkeun ku cacad dina hiji salinan gen reséptor LDL, nalika nyandak lovastatin, tingkat koléstérén dikirangan ku 30%. Ubar kasebut malah langkung efektif digabungkeun sareng résin khusus anu ngabeungkeut asam bili na nyegah panyerepan sabalikna tina peujit.
Ayeuna, statin sering dianggo pikeun nurunkeun koléstérol plasma getih. Nalika nginum obat, timbul patarosan ngeunaan efek samping anu henteu pikaresepeun. Nanging, dina hal statin, seueur efek samping, kontras, positip. Ubar ieu tiasa merangsang aliran getih, ngalereskeun plak atherosclerotic anu parantos aya (supados henteu nyimpang tina dinding pembuluh darah sareng teu ngaganggu aliran getih), ngahambat agrégasi, sareng ogé ngaleungitkeun prosés radang dina témbok pembuluh darah. Dina pasien anu nyandak statin pikeun kahiji kalina, épék ieu dibuktikeun pisan sateuacan tingkat kolesterol mimiti turun, sareng kamungkinan dihubungkeun sareng inhibisi sintésis isoprenoid. Tangtosna, henteu unggal efek tina statin mangpaat. Dina sababaraha pasien (biasana diantara anu nyandak statin digabungkeun sareng obat-obatan sanésna anu nurunkeun koléstérol), nyeri otot sareng lemah otot tiasa lumangsung, sareng sakapeung dina bentuk anu cukup kuat. Sisi épék anu sanés seueur statin ogé kadaptar, anu, untung, jarang kajadian. Dina Lolobana pasien, nyandak statin tiasa ngahambat ngembangkeun panyakit kardiovaskular. Sapertos ubar sanésna, statin kedahna ukur dianggo sakumaha disarankeun ku dokter.

Kalayan henteuna turun tina koléstérol HDL, tingkat koléstérol pisan rendah, sareng kasakit Tangier, koléstérol henteu ditangtukeun. Duanana kalainan genetik akibat tina mutasi protéin ABC1. Fraksi koléstérol HDL bébas henteu tiasa nyandak koléstérol ti sél kakurangan ABC1, sareng sél anu dikurangan ku kolesterol gancang dipiceun tina getih sareng rusak. Kadua henteuna keturunan HDL sareng tangier panyawat jarang pisan (kurang ti 100 kulawarga sareng kasakit Tangier dipikanyaho di sakuliah dunya), tapi panyakit ieu nunjukkeun peran protéin ABC1 dina ngatur tingkat plasma HDL. Kusabab tingkat HDL plasma anu lemah dina hubungan anu luhur sareng karusakan arteri koronér anu luhur, protéin ABC1 tiasa janten target anu mangpaat pikeun ubar anu ngatur pikeun ngatur tingkat HDL. ■
Hormon stéroid dibentuk ku ngabédakeun ranté tina koléstérol sareng oksidasi na.
Hiji jalma nampi sadaya stéroid hormonna tina koléstérol (Gbr. 21-45). Dua kelas hormon stéroid disintésis dina korteks adrenal: mineralkortikoid,anu ngatur panyerapan ion anorganik (Na +, C l - sareng HC O 3 -) dina ginjal, sareng glukokortikoid, anu ngabantosan ngatur gluconeogenesis sareng ngirangan réspon radang. Hormon séksina dihasilkeun dina sél lalaki réproduktif lalaki sareng awéwé sareng dina plasenta. Diantarana progesteron anu ngatur siklus réproduksi awéwé, androgen (é. téstostéron) jeung estrogen (estradiol), anu mangaruhan pamekaran karakteristik sekundér sekunder dina lalaki. Hormon stéroid gaduh pangaruh dina konsentrasi pisan rendah sahingga janten disintésis dina jumlah anu kawilang leutik. Dibandingkeun uyah bili, koléstérol saeutik dikonsumsi pikeun produksi hormon stéroid.

Gbr. 21-45. Sababaraha hormon stéroid dibentuk tina koléstérol. Struktur sababaraha sanyawa ieu dipidangkeun dina Gbr. 10-19, v. 1.

Sintésis hormon stéroid butuh jalan ngaleungitkeun sababaraha atanapi sadaya atom karbon dina "ranté sisi" tina C-17 D-ring koléstérol. Pamindahan ranté gigir aya dina mitokondria jaringan stéroid. Proses ngaleupaskeun diwangun ku hidroksilasi dua atom karbon anu naté ti ranté gigir (C-20 sareng C-22), teras ngabeungitkeun beungkeutan antawisna (Gbr. 21-46). Wangunan rupa-rupa hormon ogé kalebet ngenalkeun atom oksigén. Sadaya réaksi hidroksilasi sareng oksidasi nalika biosintésis stéparasi dikatalisasi ku oksidase-fungsi dicampur (tambahkeun ka 21-1) anu nganggo NА D PH, O 2 sareng sitokrom Biotik P-450.
Gbr. 21-46. Ngabersihan ranté gigir dina sintésis hormon stéroid. Dina sistem oksidasi ieu sareng fungsi campuran anu dioksidasi atom karbon anu padeukeut, cytochrome P-450 tindakan minangka pamawa éléktron. Ogé kalebet dina prosés nyaéta protéin anu ngangkut éléktron, adrenodoxin sareng adrenodoxin réduktase. Sistem pemisah ranté sisi ieu kapanggih dina mitokondria korteks adrenal, dimana produksi aktif stéroid lumangsung. Pregnenolone mangrupikeun prékursor pikeun sadaya hormon stéroid sanés (Gbr. 21-45).

Panengah biosintésis koléstérol aub dina sababaraha jalur métabolik anu sanés.
Salian tina peranna salaku panengah biosintésis kolésterol, isopentenyl pyrophosphate berfungsi salaku prékursor diaktipkeun dina sintésis sajumlah ageung biomolekul nu ngalaksanakeun sababaraha fungsi biologis (Gbr. 21-47). Ieu kalebet vitamin A, E, K, pigmén tutuwuhan sapertos karoten sareng ranté pholol klorofil, karét alami, seueur minyak penting (sapertos dasar seungit minyak lemon, kayu putih, kasturi), hormon serangga serangga anu ngatur métamétosis, dolichols, anu ngawula salaku pembawa larut lipid dina sintésis kompléks polisakarida, ubiquinone sareng plastoquinone - pamawa éléktron dina mitokondria sareng kloroplas. Sadaya molekul ieu isoprénoid dina struktur. Langkung ti 20,000 isoprénoid anu béda parantos kapanggih di alam, sareng ratusan énggal anu dilaporkeun unggal taun.
Gbr. 21-47. Gambar sakabéh biosintésis isoprénoid. Struktur kalolobaan produk ahir anu dibere didieu dirumuskeun dina chap. 10 (v. 1).

Prenylasi (kantétan kovalén tina isoprenoid, tempo Gbr. 27-35) mangrupikeun mékanisme umum anu protéin jangkar dina permukaan jero mémbran sél mamalia (tempo Gbr. 11-14). Dina sababaraha protéin, lipid kabeungkeut ku gugus farnesil 15-karbon, di sajabana ogé gugus geranyl 20-karbon. Dua jinis lipid ieu ngandung énzim anu béda. Mungkin réaksi prenylasi langsung protéin kana mémbran anu béda-béda gumantung kana lipid anu dipasang. Prilinasi protéin nyaéta peran penting anu penting pikeun turunan isoprene - pamilon jalur jalur metabolis koléstérol.
Ringkesan Bagian 21.4 Biosintésis Koléstérol, Stéroid, sareng Isoprénoid
■ Koléstér kabentuk tina acetyl-CoA dina urutan réaksi anu rumit ngalangkungan perantara sapertos β-hydroxy-β-methylglutaryl-CoA, mevalonate, dua isoprene dimethylallyl pyrophosphate sareng isopentenyl pyrophosphate. Kondensasi satuan isoprén masihan squalene non-siklik, anu siklus pikeun ngabentuk sistem cincin kondénsasi sareng ranté sisi stéroid.
■ Sintésis koléstérol dina kontrol hormonal,, salian, dipeungpeuk ku ningkatkeun konsentrasi kolésterol intrélular, anu lumangsung ku modifikasi kovalén sareng pangaturan transkripsi.
■ Éster kolesterol sareng kolésterol dibawa ku getih salaku lipoproteins plasma. Fraksi VLDL nransfer koléstérol, éster koléstilol sareng triacylglycerol ti ati ka jaringan séjén, dimana triacylglycerol dibersih ku lipoprotein lipase sareng VLDL dirobih kana LDL. Fraksi LDL diperkirakeun dina éster koléstérol sareng éster kololér sacara henteu langsung direbut ku reséptor ku endocytosis, sedengkeun apolipoprotein B-100 di LDL diakui ku reséptor mémbran plasma. HDL ngahapus koléstérol tina getih, mindahkeun kana ati. Kaayaan gizi atanapi cacat genetik dina metabolisme kolesterol tiasa ngakibatkeun atherosclerosis sareng infark miokard.
■ Hormon stéroid (glukokortikoid, mineralosorticoid sareng hormon seks) kabentuk tina koléstérol ku ngarobih ranté gigir sareng ngenalkeun atom oksigén kana sistem stok na cincin. Seueur sanyawa isoprénoid sanésna dikaluarkeun tina mevalonate ku kondensasi isopentenyl pyrophosphate sareng dimethylallyl pyrophosphate sapanjang sareng koléstérol.
■ Prénsiasi protéin nu tangtu ngarahkeun ka situs beungkeut séléran membran sareng penting pikeun kagiatan biologis na.
Pananya 48. Peraturan métabolisme asam lemak luhur (β-oksidasi sareng biosintésis). Sintésis malonyl CoA. Karboksilase Acetyl CoA, régulasi kagiatan na. Angkutan acil Co-a ngalangkungan mémbran mitochondria.
Utama
jumlah fénlalanin dikonsumsi
ku 2 cara:
hurung
dina bajing,
kabukti
dina tirosin
Ngahurungkeun
phenylalanine ka tirosin utamina
perlu ngaleupaskeun kaleuwihan
phenylalanine, ti saprak konsentrasi tinggi
Anu beracun ka sél. Atikan
Tirosin henteu masalah pisan
saprak kurangna asam amino ieu
dina sél sacara praktis henteu kajadian.
Utama
Métabolisme phenylalanine dimimitian
kalayan hidroksilasina (Gbr. 9-29), di
hasilna tirosin
Réaksi ieu dikandung ku anu tangtu
m soaxy-nase - phenylalanine hydra (zsilase,
anu ngagaduhan salaku produsén co-produsén
tetrahydrobiopterin (N4BP).
Aktivitas énzim gumantung kana ogé
ku ayana Fe2.
Di
haté utamana ngagancangan mobilisasi
glikogén (tingali bagian 7). Nanging saham
glikogén dina ati dileungitkeun
Dina puasa 18-24 jam puasa. Sumber utama
glukosa salaku saham amprok
glikogén janten glukoneogenesis,
anu mimiti ngagancangkeun
Gbr.
11-29. Parobihan métabolik anu utami
énergi nalika ngarobah nyerep
kaayaan postabsorbent. CT
- awak keton, FA - asam lemak.
4-6 h
saatos tuang terakhir. Substrates
gliserol dipaké pikeun sintésis glukosa,
asam amino na laktat. Dina luhur
Laju sintésis konsentrasi glukagon
asam lemak ngirangan kusabab
fosforilasi sareng aktifitas
karboksilatase sareng acetyl CoA
nambahan-oksidasi nambahan. Nanging,
tambah suplai gajih kana ati
asam anu diangkut
ti depot gajih. Acetyl-CoA kabentuk
dina oksidasi asam lemak, éta dianggo
dina ati pikeun sintésis awak keton.
Di
jaringan adipose kalawan ningkatkeun konsentrasi
laju sintésis glukagon
TAG sareng lipolysis dirangsang. Stimulasi
lipolysis - hasil aktivasina
sensitip hormon TAG lipase
adipocytes dina pangaruh glukagon.
Asam Lemak Jadi Penting
sumber énergi dina ati, otot sareng
jaringan adiposa.
Janten
sahingga, dina periode postabsorption
konsentrasi glukosa getih dijaga
dina tingkat 80-100 mg / dl, sareng tingkat lemak
asam sareng awak keton naék.
Gula
diabetes mangrupikeun panyakit anu lumangsung
kusabab mutlak atanapi dulur
kakurangan insulin.
A.
Bentuk utama klinis gula
diabetes
Numutkeun
Organisasi Dunya
diabetes kasehatan
digolongkeun dumasar kana bédana
faktor genetik sareng klinis
dua bentuk utama: diabetes
Tipe I - gumantungkeun ku insulin (IDDM), sareng diabetes
Tipe II - bebas non-insulin (NIDDM).
Peraturan
sintésis zhk. Énzim pangaturan
sintésis lcd - acetyl CoA karboksilat.
Énzim ieu diatur ku sababaraha
cara.
Aktipkeun / Dissociation
kompleks subunit énzim. Di
bentuk aktif tina acetyl CoA karboksilat
ngagambarkeun kompleks anu misah,
masing-masing diwangun ku 4 subunit.
Aktivator énzim nyaéta sitrat. Éta ngarangsang
kombinasi kompléks, salaku hasilna
dimana kagiatan énzim naék
. Inhibitor-palmitoyl-CoA. Anjeunna ngageroan
dissociation rumit sareng turun
aktivitas énzim.
Fosforilasi / Dephosphorylation
acetyl CoA karboksilat. Di
kaayaan postabsorption atanapi di
karya fisik glukagonisasi
adrenalin ngaliwatan siklus adenylate siklus
sistem diaktipkeun ku prokinase A sareng
merangsang subunit fosforilasi
acetyl CoA karboksilat. Fosforilasi
énzim teu aktif sareng sintésis lemak
asam eureun.
Nyerep
periode insulin ngaktifkeun fosfatase,
sareng karboksilatase acetyl-CoA
kaayaan dephosphorylated. Teras
dina pangaruh citrate lumangsung
polimérisasi protomér énzim, sareng
anjeunna janten aktip. Salaku aktivasina
énzim, citrate ngalaksanakeun anu sanés
fungsi dina sintésis LCD. Nyerep
période dina mitokondria sél ati
ngumpulkeun citrate, di mana
sésa acyl diangkut ka
sitosol.
Peraturan
Tingkat ox-oksidasi
Β-jalur oksidasi-métabolik,
pageuh numbu ka pagawéan CPE sareng umum
cara katabolisme. Ku sabab eta laju
diatur ku sél peryogi pikeun
tanaga i.e. ku babandingan ATP / ADP sareng NADH / NAD, ogé laju réaksi CPE sareng
jalur umum katabolisme. Laju
β-Oksidasi dina jaringan gumantung kasadiaan
substrat, i.e.
dina jumlah gajih
asam asupkeun mitokondria.
Konsentrasi Asam Lemak Apik
dina getih naék kana aktivasina
lipolysis dina jaringan adipose nalika puasa
dina pangaruh glukagon na nalika fisik
kerja na handapeun pangaruh adrenalin. Dina ieu
asam lemak janten
sumber énergi anu utami
pikeun otot sareng ati, salaku hasilna
β-Oksidasi dibentuk ku NADH sareng ngahambat acetyl-CoA
Kompleks dehidrogenase piruvat.
Transformasi ngabentuk piruvat
tina glukosa ka acetyl-CoA turun.
Metabolit panengah ngumpulkeun
glikolisis sareng, khususna, glukosa-6-fosfat.
Glukosa-6-fosfat nyegah hexokinase
sarta ku sabab kalemahan
panggunaan glukosa dina prosés éta
glikolisis. Ku alatan éta, utami
ngagunakeun lcd salaku sumber utama
énergi dina jaringan otot sareng ati
ngahemat glukosa pikeun jaringan saraf sareng
sél getih beureum.
Β-Oksidasi laju ogé
gumantung kana kagiatan énzim
seliltranferases karnitine kuring.
Dina ati, énzim ieu dipeungpeuk.
malonyl CoA, zat anu kabentuk
kalayan biosintésis lcd. Dina periode nyerep
glikolisis diaktipkeun dina ati na
formasi acetyl-CoA nambahan
ti piruvate. Réaksi sintésis munggaran
konversi lcd tina acetyl-CoA pikeun malonyl-CoA.
Malonyl-CoA nyegah id-oksidasi lcd,
anu tiasa dianggo pikeun sintésis
gajih
Atikan
malonyl-CoA ti acetyl-CoA-pangaturan
réaksi dina biosintésis lcd. Réaksi mimitina
konversi lcd konvérsi acetyl-CoA ka
malonyl CoA. Énzim katalitik
réaksi ieu (acetyl Coa carboxylase),
kagolong kana kelas ligan. Anjeunna ngeusian
beungkeut biotin kabeungkeut. Dina mimitina
tahapan réaksi kovalén
meungkeut biotin kusabab énergi
ATP, dina tahap 2 COO- dialihkeun
dina acetyl-CoA pikeun ngabentuk malonyl-CoA.
Acetyl CoA Carboxylase Énzim Aktivitas
nangtoskeun kagancangan sadayana sakalian
réaksi sintésis lc
citrate ngaktifkeun énzim dina sitokol
acetyl CoA karboksilat. Malonyl CoA di
kahareupna nyegah pangiriman langkung luhur
asam lemak tina sitosol dugi matriks
kagiatan nyerep mitokondria
acetyl éksternal CoA: karnitine acyltransferase,
sahingga mareuman oksidasi anu langkung luhur
asam lemak.
Acetyl-CoA Oxaloacetate →
HS-CoA Citrate

HSCOA ATP Citrate → Acetyl-CoA ADP Pi Oxaloacetate
Acetyl-CoA
dina sitoplasma ngagaduhan salaku substrat awal
dina sintésis lcd, sareng oxaloacetate di
sitosol ngalaman parobahan dina
hasil anu pyruvate dibentuk.
Biosintésis kolesterol
Biosintésis koléstérol lumangsung dina reticulum endoplasmic. Sumber sadaya atom karbon dina molekul nyaéta acetyl-SCoA, nu asalna di dieu tina mitokondria salaku bagian citrate, sakumaha sintésis asam lemak. Biosintésis koléron meakeun 18 molekul ATP sareng 13 molekul NADPH.
Wangunan koléstérol lumangsung dina leuwih ti 30 réaksi, anu tiasa dikelompokkeun dina sababaraha hambalan.
1. Sintésis asam mevalonik
Dua réaksi sintésis munggaran saluyu sareng réaksi ketogenesis, tapi saatos sintésis 3-hydroxy-3-methylglutaryl-ScoA, énzim ngalebetkeun hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA réduktase), ngabentuk asam mevalonik.
 |